盲鳗基因组揭示脊椎动物进化奥秘

编译:微科盟-麦克,编辑:微科盟悄咪咪、江舜尧。
微科盟原创微文,欢迎转发转载,本文来源《基因组Genome》公众号。
导读
作为唯一幸存的无颌鱼类谱系,盲鳗(hagfishes)和七鳃鳗(lampreys)为研究早期脊椎动物的进化提供了关键的视角。本研究以褐盲鳗(Eptatretus atami)的染色体尺度基因组序列为基础,研究了脊椎动物全基因组复制(genome-wide duplication) 和程序化DNA消除(programmed DNA elimination) 的复杂历史、时间和功能作用。结合系统发育分析的证据,本研究建立了一个完整的脊椎动物基因组进化图谱,包括早寒武纪(the Early Cambrian) cyclostome-gnathostome分裂前的同源四倍体化(1RV),随后是中晚寒武纪的异源四倍体化(2RJV)和长期寒武纪-奥陶纪(Cambrian-Ordovician)的六倍体化(2RCY)。随后,盲鳗经历了广泛的基因组变化,染色体融合并伴随着重要器官系统基因的丧失(眼睛、破骨细胞【osteoclasts】),部分解释了它们身体结构的简化。最后,表征了盲鳗的程序性DNA消除,鉴定了发育早期从体细胞谱系中删除的蛋白质编码基因和重复元件。这些生殖系特异性基因的消除提供了一种机制,通过抑制生殖系/多能性功能来解决体细胞和生殖系之间的遗传冲突,这与在七鳃鳗中的发现相似。脊椎动物早期基因组史的重建为进一步探索脊椎动物的进化提供了一个有价值的框架。
论文ID
原名:The hagfish genome and the evolution of vertebrates
译名:盲鳗基因组与脊椎动物的进化
期刊:Nature
IF:64.8
发表时间:2024.1
通讯作者:Ferdinand Marlétaz、 Daniel S. Rokhsar、Jeramiah J. Smith
通讯作者单位:日本冲绳科学技术大学院大学、美国肯塔基大学
DOI: 10.1038/s41586-024-07070-3
实验设计
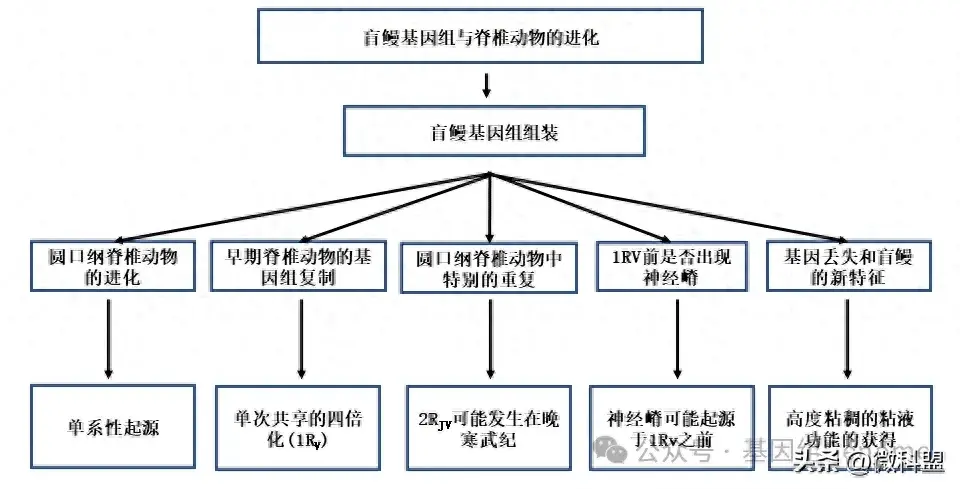
结果
1 圆口纲脊椎动物(cyclostome)的进化
利用来自睾丸的长、短reads组合对褐盲鳗(Paramyxine atami)的生殖系基因组进行了测序,并利用体细胞组织的数据将其组装成染色体级别的基因组。褐盲鳗组装约2.52 Gb,包括17个大型染色体级别的scaffolds。与预期的体细胞核型(2n = 34)一致。组装的基因组长度介于基于评估的荧光的somatic细胞(2.01 Gb)和germline细胞 (3.37 Gb),与k-mer估计的大小一致(分别为2.02和3.28 Gb)。 E. atami germline基因组还包括7条高度重复的染色体,这些染色体在发育过程中被完全消除,其序列在组装中以亚染色体片段的形式存在,类似地,在高度重复的生殖系中也发现了特定的染色体,如七鳃鳗和鸣禽(songbirds)。 总共注释了28,469个基因,其中22,663个与其他物种的蛋白质编码相似。
使用盲鳗基因集来测试cyclostomes的单系性,通过引入(i)更广泛的cyclostomes分类样本(包括大西洋盲鳗Myxine glutinosa的新数据)和(ii)改进的位点异质性(site heterogeneity)和组成偏差建模(compositional bias),扩展了开创性的早期研究。通过完整的盲鳗和七鳃鳗基因组,得到了一套新的1,467个同源基因序列,且包含比早期研究多8倍的标记。这些分析通过分区分析和位点异质模型分析(图1b)证实了cyclostomes起源的单一性。通过后验预测检验验证的六类氨基酸重编码(six-category amino-acid recoding)进一步支持了对成分异质性的稳健性。
尽管盲鳗和七鳃鳗的核型不同,但它们的染色体是相关的(图1c,d)。 经过~457 Ma的独立进化,基因顺序被高度打乱,重复的区域也是不同的。一般来说,每条盲鳗染色体通常是2到6条七鳃鳗染色体的融合,相反,每条七鳃鳗的染色体通常与一条盲鳗染色体相关,很少有例外。为了区分盲鳗谱系中可能的染色体融合和/或七鳃鳗谱系中的分裂(或重复),使用了先前重建的祖先脊索动物连锁群(clg,chordate linkage groups: A1, A2, B-Q)。虽然七鳃鳗染色体通常来自单个clg(与先前对七鳃鳗基因组的分析一致),但盲鳗染色体显然是通过来自这些祖先元件的不可逆融合。类似于但不同于在gnathostome主干谱系上观察到的融合(图1c)。盲鳗和七鳃鳗之间直接的、基本上是一对一的片段对应关系与先前的假设一致,即两种环口动物具有相同的复制历史,但是需要更详细的系统发育分析来排除趋同复制的可能性。
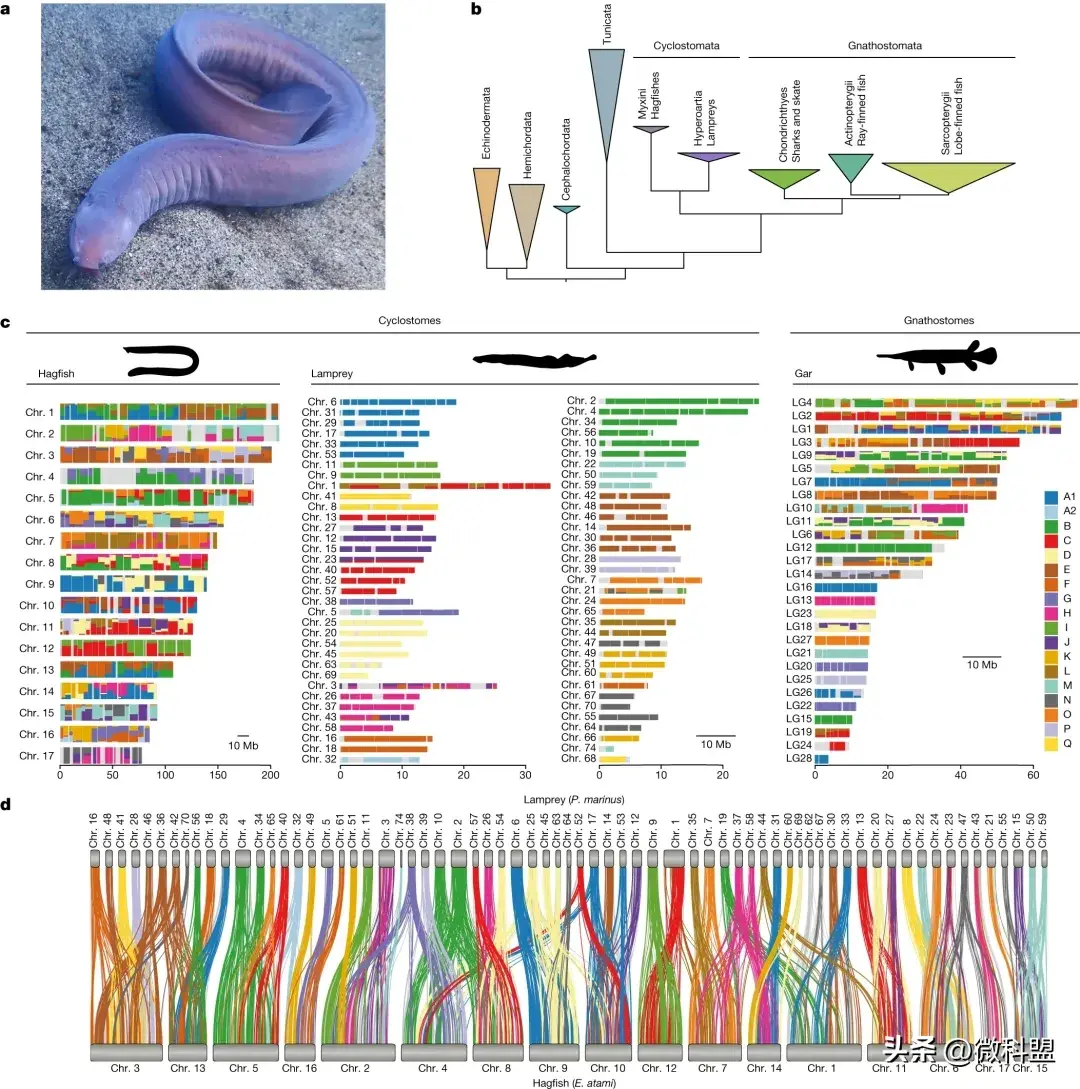
图 1. Cyclostome和gnathostome的系统发育关系和同源结构。 a,盲鳗, Eptatretus atami。b,基于选定基因使用site异构模型 (CAT+GTR)的系统发育分析。c, 核图显示盲鳗、七鳃鳗和雀鳝染色体在脊索动物祖先中连锁组的分布(CLG A1、A2、B-Q)。计算来自具有显着富集的 CLG 的基因(Fisher 检验)。盲鳗,七鳃鳗和从 Phylopic下载的雀鳝图。d,保守的共线区表明盲鳗染色体通常是多条七鳃鳗染色体的融合。连接直系同源基因的线系为根据祖先脊索动物连锁群着色(图C中的颜色图例)
2 早期脊椎动物的基因组复制
通过使用了两种互补的系统发育方法来解决早期脊椎动物序列的多倍体化事件:(i)基于模型的多倍体化推断,来自大量的基因树和(ii)基于染色体尺度共线性的具有相似进化历史的串联基因。首先测试了基于8,931个基因树的全基因组重复序列的可能假说,通过使用WHALE对基因和物种树进行概率计算。该分析支持对脊椎动物主谱系(1RV)中单基因组重复发生事件,随后是gnathostome (2RJV)和cyclostome (2RCY)系的独立多倍化(所有贝叶斯因子BFNull_vs_WGD < 10-3)(图2a)。分析中没有发现脊椎动物谱系(2RV)的第二轮多倍体化的支持,也没有发现个七鳃鳗或盲鳗谱系特有的多倍体化事件(图2a),与基于共线性的分析一致。
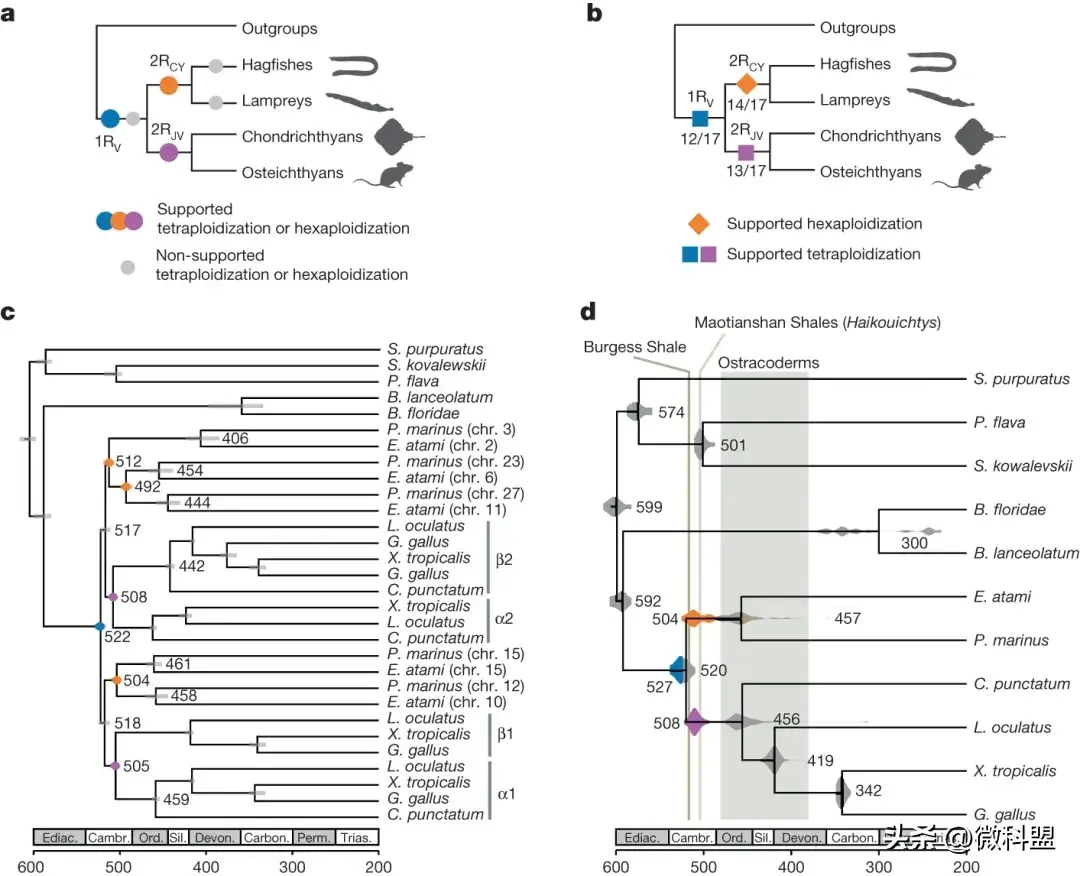
图2.脊椎动物基因组复制的历史。a, 基于基因树的早期脊椎动物进化中多倍体化事件的概率推断(WHALE)支持所有脊椎动物共有的初始四倍体化(1RV)、gnathostome特异性四倍体化(2RJV)和cyclostome 特异性多倍体化(2RCY)。支持的多倍体化事件(贝叶斯因子 BFNull_vs_WGD < 10-3)以颜色(1RV、2RJV、2RCY)显示,不支持的多倍体化事件以灰色(2RV、盲鳗特异性、七鳃鳗特异性)显示。这里,WHALE方法无法区分四倍体化和六倍体化事件。b,基于旁系同源子的多倍体化推断,使用为17个CLG中的每一个重建的分子构建系统发育。脊椎动物进化过程中的连续多倍体化事件显示为彩色多边形,显示这些重复节点的CLG树的比例如图所示。c, CLGJ的样例旁系同源树。至于基因树,在旁系同源树中,一些节点对应于物种形成事件(灰色),而另一些节点对应于重复事件(彩色);这两种类型的事件都可以使用分子钟进行测算。d,分子测年早期脊椎动物进化中的多倍体化和物种形成事件。表示物种形成(灰色)和重复(a中颜色)节点的发散时间。在 c 和 d 中,每个节点都用均值标记CLG 之间的发散时间。
作者还开发了一种基于synteny-based的方法,利用有共同进化史的连锁基因群来增强单个基因树有限的系统发育信息,并避免差异基因丢失所产生的混淆效应。在这种方法中,通过其在盲鳗、七鳃鳗和几种有gnathostome中的同源基因的溯源,来确定的CLG的重复历史(例如,图1d中相同颜色的染色体或染色体片段)。在每个物种中,旁系染色体片段被称为“旁系染色体”。由于clg保存在不同的无脊椎动物中,相应的染色体连锁基因可以连接起来,为系统发育和分子定年提供外群。因此,基于18个旁系的分子系统发育关系,重建了17个CLGs/原始脊椎动物染色体;第18组包含相对较少的跨脊椎动物分类群一致关联的基因,在七鳃鳗和颌口动物中都具有异常特性.
基于旁系基因的分子系统发育支持在cyclostome-gnathostome分裂之前的单个早期脊椎动物自四倍体化(1RV),随后是后来的gnathostome特异性异源四倍体化(2RJV)和cyclostome特异性多倍体化(2RCY)(例如图2b中的CLGJ)。在14个CLG系统发育中,有12个的1RV重复节点先于cyclostome-gnathostome物种形成,并且bootstrap支持BP>60。因此,脊椎动物系统上单次共享的四倍化(1RV)与来自单基因树和基于旁系同源的系统发育的基因组重复的概率推断是一致的。分子测年表明,重复和物种形成事件发生在紧密的演替过程中,估计1RV类群的分化发生在~527亿年前,而cyclostome-gnathostome分裂发生在~520亿年前。
对1RV平行分化的估计日期对应于同源重组(再倍化)的停止,而不是整个基因组复制本身,如最初在参考文献中所指出的那样。通过比较先前提出的祖先和谱系特异性再倍化模型下基因树的可能性测试了clg谱系特异性再倍化(相对于cyclostome-gnathostome分化)(图3a)。作者发现,所有clg在1RV后的祖先再倍化得到了更大数量的显著基因树的支持(图3b),这表明减数分裂再倍化在cyclostome-gnathostome分裂时基本上完成了。这与其他最近的脊椎动物自多倍体形成对比,其中许多同源染色体通过随后的物种形成事件保持了四体遗传。不幸的是,分子系统遗传学只能估计1RV同源四倍体化事件本身的时间,因为它可能被持续时间未知的同源重组所掩盖:1RV复制事件可能比1RV同源物的分化时间(约527亿年前)早数百万年。

图3.脊椎动物基因组复制后的有限谱系特异性再二倍体化。a,1RV后祖先再二倍体化(左)和谱系特异性再二倍体(右)模型下的基因树拓扑结构。在祖先再二倍体化设想中,旁系同源基因序列在cyclostome- gnathostome 分裂之前分化,从而通过复制的基因拷贝进行分组。在谱系特异性再二倍体情景中,旁系同源序列在gnathostome 和cyclostome谱系中独立发散,因此基因按谱系分组。b,对于 17 个信息性祖先连锁组 (CLG) 中的每一个,支持 1RV后祖先和谱系特异性再二倍化情景的显着支持的基因树数量。c,2RCY后祖先和谱系特异性再二倍体模型下的树拓扑结构。CLG B 旁系同源树显示了1RV copy的祖先再二倍体化拓扑,其中两个盲鳗(chr4和 5)和两个七鳃鳗(chr10和 2)旁系同源独立再二倍体化。d,由CLG B旁系同源系统发育解析的脊椎动物Hox基因簇的进化史(见图c底部)。
3 圆口纲脊椎动物(cyclostome)中特别的重复
基于旁系同源基因的分子系统发育也有力地支持和完善了2RJV四异源倍体化的设想。旁系同源树的分子定年表明,前2RJV α - β祖细胞的分裂发生在中寒武纪~508 Mya(图2)。然而,异源四倍体化事件本身(即α和β祖细胞的杂交和随后相关的基因组加倍)发生在α - β分化之后的一段时间,而且本身不能用分子系统发育来精确定位。在最近的异源四倍体脊椎动物中,如Xenopus和金鱼,杂交发生在祖先分化的10-15百万年内。如果研究人员将其作为齿颌异体四倍体的类比物,那么2RJV可能发生在晚寒武纪,远早于gnathostomes的起源~456 Mya,接近中晚奥陶世边界。
在圆口纲脊椎动物中,旁系同源树证实了盲鳗和七鳃鳗染色体片段的同源性(图2c)。通常在每个CLG上观察到一个或两个重复节点,表明在盲鳗和七鳃鳗分裂前发生了共享的圆口纲脊椎动物全基因组复制~457亿年前。
同源(即旁系)圆口纲脊椎动物染色体(峰值在~511和~493)Mya处的divergence次数双峰分布与在鲟鱼中通过二倍体和相关四倍体杂交观察到的两步六倍体化事件一致。在正式测试了谱系特异性再倍化(图3c),发现尽管盲鳗和七鳃鳗染色体片段同源物之间接近一对一的关系(图1d),但2RCY后的再倍化在很大程度上是由crown圆口纲脊椎动物完成的。根据估计的旁系同源分化时间和旁系同源树拓扑结构,确定了一个谱系特异性再倍化的案例,在2RCY后影响了盲鳗和七鳃鳗的CLGB旁系同源物。具体来说,从CLGB的1RV拷贝继承下来的盲鳗chr4/chr8和七鳃鳗chr10/chr12的旁系同源物对分别在盲鳗和七鳃鳗中独立地进行了再倍化,这可以通过CLGB旁系同源系统发育(图3c)和估计的旁系同源分化(圆口纲脊椎动物分裂~457 Ma,盲鳗chr4 - chr8旁系同源分化~431 Ma,七鳃鳗chr10 - chr2 旁系同源分化~442 Ma)来证明。
4 脊椎动物Hox群的进化
值得注意的是,CLGB包含Hox簇,这是脊椎动物WGD(whole genome duplication)早期分析中的一个关键位点。通过CLGB 旁系和Hox-plus-bystander树,完全解决了在1RV, 2RCY和2RJV事件之后的四个gnathostome和六个cyclostome Hox集群的进化史(图3d)。 虽然以前的研究未能确定七鳃鳗和盲鳗Hox簇之间的一对一关系,但作者报告了七鳃鳗Hox ζ, α, γ, δ与盲鳗Hox II, III, IV, VI之间明确的同源关系,分组如下:ζ-II, α-III, γ-IV和δ-VI。相比之下,在CLGB的谱系特异性再倍化之后,七鳃鳗Hox β/ε和盲鳗Hox V/I之间不存在真正的(即一对一)同源关系。
5 神经嵴起源
基于旁系同源基因的分类使研究人员能够将旁系准确地分配给特定的重复,这揭示了脊椎动物特征(如神经嵴、基板【placodes】、或激素系统【placodes】)的相对起源。正如Wada等人最初指出的那样,两个旁系分支是否保留(或分割)祖先成分可以帮助确定一个片段可能是在1RV之前还是之后出现。
为了评估神经嵴的起源,作者考虑了一组涉及神经嵴specification和migration的22个基因家族(图4a),并根据文献和现有RNA-seq数据,整理了相应的1RV序列是否在gnathostome和七鳃鳗中执行ncc相关功能。作者发现,在这些基因家族中,包括Tfap2、SoxE、EdnR、Twist1和Gata3,两个1Rv分支上的同源基因都参与神经嵴相关功能。这一模式表明,ncc相关功能可能遗传自脊椎动物祖先的1Rv前的基因,因此表明神经嵴可能起源于1Rv之前。wgd后的亚功能化在其出现中发挥了有限的作用,这与其他gnathostome的新特征(如肢体【limbs】)相反。一致地,在七鳃鳗中,另一种1R序列似乎也参与NCC,例如Gata3, six1, Msx1和可能的FoxD(图4a)。
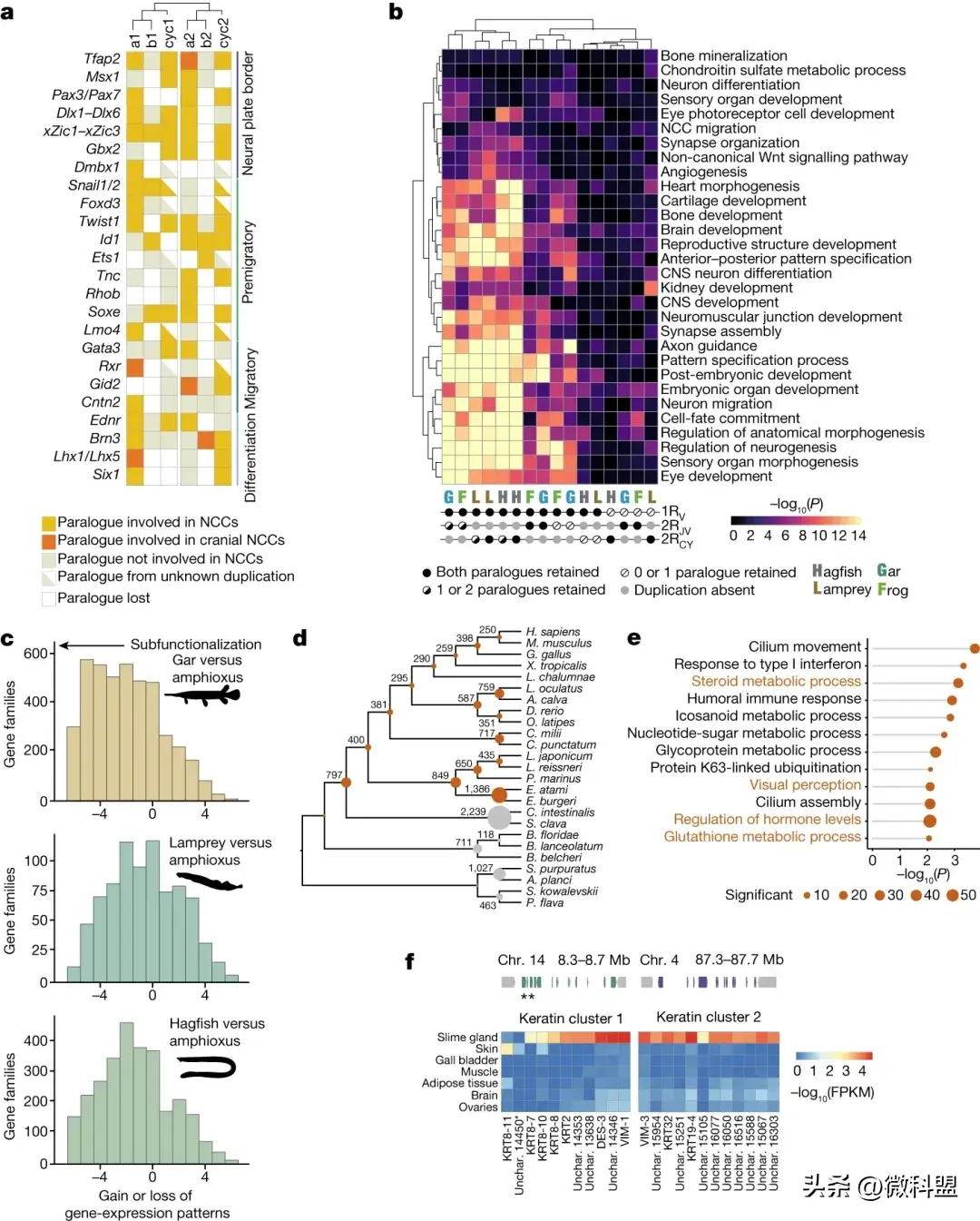
图4.脊椎动物WGD的功能影响和脊椎动物基因丢失。a,关键的神经嵴相关基因家族,其成员根据其相对于 1RV 和 2RJV 的功能作用(颜色)和旁系状态进行分类。在gnathostome和七鳃鳗中,来自 1RV 的两个拷贝的旁系同源物参与 NCC 相关功能,支持了 NCC 早于 1RV 的假设。b,在脊椎动物WGDs之后显示特定保留模式的基因集中的功能注释术语(Go ontology;GO)的富集。每列对应一组旁系同源基因特定物种中复制后保留的特定模式。c,特定脊椎动物之间阳性器官特异性表达域差异的分布. d,后口动物(deuterostomes)的基因家族突出了盲鳗谱系相对于其他脊椎动物和后口动物(灰色)的严重丢失。e,盲鳗谱系中丢失的基因家族的功能富集(GO),突出了视觉和激素系统的简化(橙色标签)。f,14号和4号染色体上两个α-角蛋白基因簇的结构,以及它们在粘液腺和皮肤中的表达,如热图所示。基因在热图中的显示顺序与它们位于两个簇中的顺序相同。星星表示两个基因优先在皮肤中表达。
相比之下,在cyclostome、骨鱼目动物(osteichthyans)甚至羊膜脊椎动物动物(amniotes)中,躯干(trunk )和颅骨(cranial)NCCs的建立似乎有所不同,涉及不同的基因。参与这一过程的一些基因(但不是全部)似乎显示出最近发生的亚功能化。例如,参与颌骨颅NCC的Lhx5、Id3或Gid2或Dmbx没有具有类似功能的1RV或2RJV旁系基因,而参与颅骨和主干NCC祖先规范的Tfap2和sox212在参与该功能的1RV分支上都有旁系基因。七鳃鳗表现出明显的差异:在七鳃鳗migration神经嵴细胞中,RhoB和Ets均未参与或表达,而Lhx5、Dmbx和Ets1基因在后来的NCC衍生物中均有表达。尽管盲鳗谱系经历了广泛的基因丢失(见下文),但作者找回了研究的大多数ncc相关同源基因。需要进一步的功能研究来确定盲鳗中后续的2RCY同源基因是否被纳入该谱系特有的ncc相关功能。
6 旁系同源基因---截然不同的命运
经过两轮基因组复制后,cyclostome中保留的旁系同源基因在功能上与发育和神经系统活动的调节有关。为了确定相似基因在1RV之前/在cyclostome特异性2RCY后是否优先保留在圆口纲脊椎动物谱系的多个拷贝中,测试了显示功能富集不同保留模式的旁系集(图4b)。复原了之前发现的gnathostomes旁系富集的GO(例如,轴突引导,胚胎器官发育),但有趣的是,作者发现它们优先与泛脊椎动物1RV后保留的旁系相关,而不是和它们在之后的gnathostomes重复2RJV相关(图4b),这表明1RV在脊椎动物神经系统的早期发育中起关键作用。然而,在cyclostomes 中,这些GO terms优先与所有多倍体(1RV和2RCY)后系统保留的旁系同源物联系在一起,这表明在功能水平上,旁系同源物的进化路径不同,可能与2RCY后的保留比2RJV增加有关。
在WGD后,旁系同源基因的命运通常与它们获得更具体的表达域有关,这可以解释亚功能化和功能创新。为了研究cyclostomes-gnathostomes动物的基因表达差异模式,比较了文昌鱼、雀鳝、七鳃鳗和盲鳗在6个一致器官中的旁系同源基因。考虑到3,009个基因家族,发现基因在雀鳝中的表达特异性高于七鳃鳗和盲鳗,而盲鳗的特异性最低。然后,计算了文昌鱼和不同脊椎动物物种在同一基因家族中获得或丢失的表达模式的数量,同样表明种cyclostomes的亚功能化水平低于gnathostomes(图4c)。最后,使用基因表达聚类(WGCNA,)显示出显著的旁系同源基因富集。有趣的是,发现只有神经组织在gnathostomes鱼(如gar)和盲鳗中都表现出富集,而许多最近复制的基因以器官特异性的方式表达。综上所述,这些结果表明,与gnathostomes,cyclostomes更大程度上,比如盲鳗,表现出更有限的功能化或表达模式的特化。
7 基因丢失和盲鳗的新特征
盲鳗在脊椎动物中丢失的基因最为广泛,共有1386个基因家族缺失,其中 892个基因家族存在于后口动物(deuterostome)祖先中(图4d)。盲鳗突出表现为失去了几个完整基因家族的所有成员,而不仅是旁系同源基因数量的丢失。
盲鳗中丢失的几个基因家族在功能上富集了与盲鳗缺失性状相关的功能(图4e)。例如,构成脊椎动物眼睛晶状体的γ -晶体蛋白(gamma-crystallins)在只在盲鳗中缺失(而在七鳃鳗和颌口鱼中独立存在),参与光感受器维持和发育的EYS (eyes Shut homologue)和RBP3(gamma-crystallins)基因也缺失。有趣的是,在其他脊椎动物中参与骨发育及其激素控制的几个基因在盲鳗中缺失:控制gnathostomes破骨细胞增殖的RANK/Osteoprotegerin通路的两个成员以及编码甲状旁腺激素(PTH和256 PTLH)的基因参与钙代谢的调节(它们的受体仍然存在)。这些基因存在于七鳃鳗中,它们在盲鳗谱系中的缺失可能与盲鳗椎软骨的有限凝结有关。
盲鳗也获得了新的特征,最引人注目的是它们分泌一种高度粘稠的粘液(slime)的惊人能力,这种粘液在躲避捕食者时起着重要作用。作者发现在黏液腺体中特异性和高表达的两个基因簇与中间细丝(α -角蛋白)相关。有趣的是,这个簇中有一个包含主要在皮肤而不是粘液腺中表达的基因,这与最近的文献的观点一致,盲鳗粘液的角蛋白线可能起源于皮肤的组成部分(图4f)。作者发现黏液腺中表达最多的糖蛋白包括血管性血液病A和D结构域,而不是之前假设的黏液蛋白型结构域。
8 程序化DNA和基因消除
盲鳗的体细胞(somatic)和生殖细胞(germline)表现出不同的核型,这显然是由于胚胎程序化DNA消除导致种系特定染色体的丢失。根据k-mer计数,估计约3.3 Gb E. atami 生殖系基因组中约1.3 Gb丢失,与细胞荧光测定结果一致。基因组组装分析鉴定了大量胚系特异性基因,证实胚系特异性区域含有大量复杂的重复元件,包括一个新鉴定的重复,占基因组的4%(图5)。
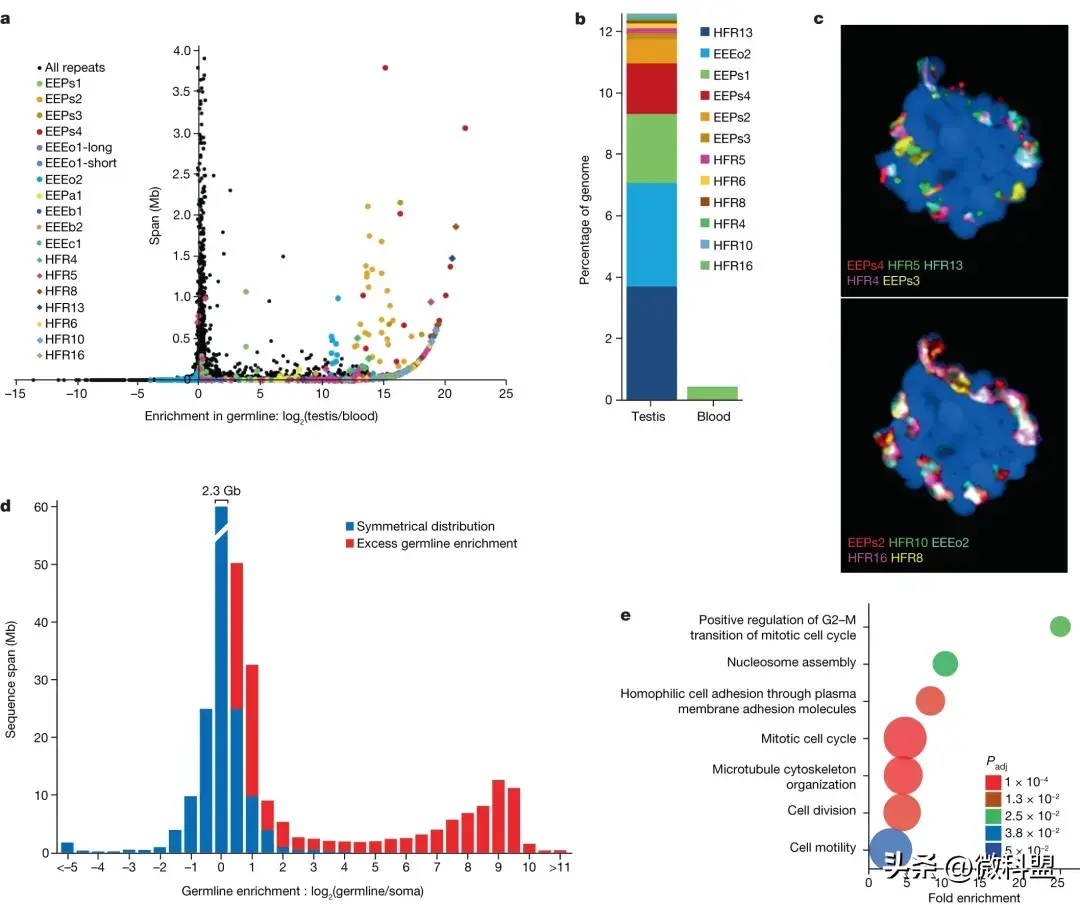
图5.盲鳗中的种系特异性/富集序列和基因。a,胚系富集图以及E. atami基因组中所有预测重复元件的估计跨度,重点关注累积跨度为 <4Mb(每个家族成员)的元件。先前确定的元素用彩色圆圈突出显示,新识别的复制元素用彩色菱形突出显示。c,FISH将高拷贝胚系特异性重复序列杂交到睾丸中期平板上,显示它们在染色体内独特的空间聚类(蓝色复染是NucBlue:Hoechst。d,比较从胚系(睾丸)与体细胞(血液)组织中提取的DNA的序列深度,确定了大量的基因组区间,并有证据表明胚系中高度富集。e,在胚系特异性区域内编码的基因富集了与细胞周期和细胞运动调节相关的Go terms.
到目前为止,还没有在任何盲鳗物种中发现生殖系特异性基因。通过比较组装基因组的低拷贝区和中拷贝区生殖系(胚)系和体细胞的reads深度,鉴定了 E. atami 的生殖系特异性基因(图5 d)。发现了81 Mb的生殖系特异性序列,编码1654个基因,其中226个具有可识别的人类同源基因(121个非冗余的人类基因)。46个种系特异性间隔中有44个可以pcr扩增,来自睾丸,但不能扩增血液DNA(验证率为95.7%)。基于GO分析,盲鳗的生殖系特异性基因具有多种生物学功能,包括与细胞周期、细胞运动和染色质/DNA修复相关的功能(图5e)。在七鳃鳗的生殖系特异性基因中也富集了类似的功能,这支持了在体内被消除的基因通常具有有利于种系发育和维持的功能的假设。
盲鳗和七鳃鳗生殖系特异性基因在功能上的广泛相似性表明,DNA消除可能是cyclostome谱系共同的祖先特征。为了试图确定理论祖先生殖系特异性染色体的痕迹,作者检查了被淘汰基因的直系同源和旁系同源类群。尽管盲鳗和七鳃鳗被淘汰的基因在功能上普遍相似,但在两个基因组中都发现了很少的同源基因被淘汰。在非冗余盲鳗基因家族中,共有7个在七鳃鳗中被剔除(CDH1/2/4、GJC1、MSH4、NCAM1、290 SEMA4B/C、WNT5/7A/B和YTHDC2。对基因树的检查表明,其中三个(MSH4、WNT7A和YTHDC2的同源物)共享一个最后的共同祖先,该祖先可以追溯到基础脊椎动物分化/复制事件之后的单一谱系。这一小组基因可能反映了共享的生殖系特异性序列的痕迹,这些序列在cyclostome谱系早期被消除了,或者,这些基因可能在两个谱系的早期进化中被独立地招募到生殖系特异性部分。
鸣禽(songbirds)和七鳃鳗的生殖系特异性染色体不断捕获体细胞基因复制,建立新的生殖系特异性基因,由于其独特的选择性和突变性基因组环境,这些基因通常会迅速进化。在 E. atami中,观察到几个生殖系特异性基因在复制/易位到生殖系染色体后经历了额外的复制。生殖系-特异性拷贝数最高的基因是FBXL4的同源基因,FBXL4是E3泛素连接酶的调节剂,调节组蛋白去甲基化酶KDM4A68的蛋白酶体-转换(25拷贝)和TRRAP, TRRAP是几种组蛋白乙酰转移酶复合物的组成部分(18拷贝)。在近亲盲鳗E. burgeri的基因组草图中,fbxl4同源物也包含45个旁系同源物,这表明生殖系特异性FBXL4的起源和该基因家族的扩展可能早于这两个盲鳗物种之间的分裂,在过去的几百万年里,额外的谱系特异性扩展/缺失导致了旁系同源物数量的差异。这些基因家族似乎在最近经历了大量的扩展,甚至在最近的过去,强调了它们的高周转率。
表观遗传沉默标记的积累和调控降解与七鳃鳗生殖系系特异性染色体消除的细胞机制有关。这表明盲鳗DNA消除机制的组成部分可能由种系特异性染色体本身编码,或者在盲鳗生殖细胞发育的其他方面起作用。与FBXL4和TRRAP相同通路的其他基因也在E. atami生殖系特异性染色体中复制,尽管复制程度较低。这些包括:KLHL10,参与精子发生的E3泛素连接酶复合物的组成部分(4拷贝),SIN3,一种转录抑制因子,其人类同源物在睾丸中高度表达(4拷贝),DNMT1,负责在DNA复制后维持DNA 甲基化标记沉默的主要酶(2拷贝)。重要的是,这五个生殖系特异性基因家族中的每一个都至少拥有一个体细胞保留的旁系,这表明与泛素化和染色质状态调节相关的基因家族的种系特异性扩增是在大致完整的祖先体细胞途径的背景下进化的。
讨论
早期脊椎动物的进化伴随着一系列古老的多倍体化事件,这些事件很难用传统的基于序列的分子系统发育学来明确地解决。作者使用hagfish基因组和一种新的方法,将重点放在染色体规模的系统发育上,以完全解决这一古老脊椎动物多倍体的历史(图2)。最早的重复,1RV,发生在早寒武纪(~527 Ma),比最早的脊椎动物化石Haikouichthys和Myllokunmingia出现早~10 Ma。事件发生时间的相似性是巧合还是因果关系还有待观察。
在共享复制之后,在寒武纪晚期至奥陶纪早期的年间,cyclostomes-gnathostomes经历了独立的多倍体化,与脊椎动物化石记录的空白相吻合。然而,研究人员可以通过将基因复制的当代功能与其在特定复制事件中的出现联系起来,开始将早期基因组事件与脊椎动物新特征的出现和发展联系起来。例如,当一个基因在神经嵴中起作用时,它的1RV类似物也会起作用,如果神经嵴调节回路在1RV之前已经存在,那么就可以预期。更值得推测的是,研究人员注意到Evx同源基因,其中在配对鳍和四肢的发育和模式中起作用,在1RV时复制,在gnathostomes中保留了谱系(Evx1-HoxA和Evx2-HoxD),但由于谱系特异性缺失,cyclostomes缺失了HoxC/ d相关的Evx 旁系基因。这一观察结果表明,1RV重复序列可能在啮齿动物谱系进化的早期就在鳍芽的发育和模式形成中发挥了作用,这与在早期分化的galeaspids中对成对鳍襞形态的观察结果一致。
最后,与其他脊椎动物相比,对E. atami 生殖系特异性染色体的分析支持以前的假说,即这些染色体编码有利于生殖细胞发育和配子产生的功能,并表明生殖系特异性基因含量的快速更替可能是高度分化谱系中共同的特征。与其生物学的其他特征一样,七鳃鳗和盲鳗种系特异性染色体基因含量的差异可能反映了它们独立进化的漫长历史,以及自最后一个共同的cyclostomes祖先以来约4.6亿年里它们在生殖、生态和发育生物学方面积累的巨大差异。




















 鲁公网安备37020202370234号
鲁公网安备37020202370234号