金鱼端脑细胞类型图谱揭秘:空间结构与细胞进化多样性探索

编译:微科盟 Lucky,编辑:微科盟 景行、江舜尧。
微科盟原创微文,欢迎转发转载,转载请注明来源于《转录组》公众号。
需要原文请私信获取!
导读
硬骨鱼是脊椎动物中最大的类群,表现出多种多样的适应性行为。为了研究硬骨鱼类多种多样的适应行为的神经基础,本研究对金鱼端脑进行了系统的比较研究。研究者利用单细胞RNA测序和空间转录组学绘制细胞类型图谱,从而获得全新的分子神经解剖学划分。小鼠和金鱼的神经胶质细胞在4.5亿年的进化过程中高度保守,而神经元表现出基因表达的多样性和模块化。具体来说,生长抑素中间神经元(哺乳动物同型皮质中用于局部抑制性输入的神经元)聚集在一个单一的金鱼端脑核中,但在分子上是保守的,包括纹状体(纹状体是羊膜动物动机行为的中枢)在内的大脑核团在分子上与金鱼具有同源性。本研究描述了金鱼和斑马鱼在神经元分类系统之间的分子相似性,为进一步了解远古脊椎动物的外翻端脑奠定了基础。
论文ID
原名:A telencephalon cell type atlas for goldfish reveals diversity in the evolution of spatial structure and cell types
译名:金鱼端脑细胞类型图谱揭示空间结构和细胞类型进化的多样性
期刊:SCIENCE ADVANCES
IF:13.6
发表时间:2023年11月
通讯作者:Ronen Segev,Amit Zeisel
通讯作者单位:以色列理工学院
DOI号:10.1126/sciadv.adh7693
实验设计

结果
1 空间分辨率的金鱼端脑细胞类型图谱
单细胞RNA测序(scRNA-seq)能够完全基于全基因组基因表达谱(转录组)来比较不同物种间特定组织细胞类型的分子相似性和差异。而空间转录组(ST)可以保留mRNA分子在组织中的位置信息,并实现基因谱的空间定位。本研究结合这两种技术,生成了金鱼端脑的空间注释细胞类型图谱(图1A)。
首先,研究者对完整的金鱼端脑进行scRNA-seq。从10条成年期金鱼的新鲜冠状切片中,显微解剖金鱼端脑的外侧、内侧、腹侧和前侧(图1B和图S1)。每个区域都被分离成高度存活的细胞悬浮进行测序,并映射到参考基因组。由于金鱼转录组缺乏注释,研究者根据小鼠的同源基因识别并注释。如图1所示,根据细胞的表达谱将细胞划分为假定的细胞类型。硬骨鱼祖先的基因组经历了四次全基因组复制,产生了金鱼基因旁系同源物。然而,金鱼基因旁系同源物在聚类上没有差异(图S2)。因此,金鱼基因旁系同源物被折叠,研究者根据小鼠的直系同源基因使用基因名称(例如,GAD1a和GAD1b变成GAD1)。
首先,研究者根据已知的标记物(例如神经递质转运蛋白SLC17A7、SLC17A6和SLC32A1)将约40000个细胞分为三个主要类别:谷氨酸能神经元(约23400个细胞)、GABA能神经元(约8300个细胞)和非神经元细胞(约8500个细胞)(图1C)。正如预期的那样,金鱼端脑解剖部位(外侧、腹侧、内侧和前部)在细胞类型组成方面有所不同(图1D和E以及图S1);例如,腹侧端脑(大脑皮层下)中主要为GABA能细胞。一般细胞类别包含106种明确的转录组细胞类型(图1F),其中40种是GABA能细胞,48种是谷氨酸能细胞,18种是非神经元细胞(即神经胶质细胞和免疫细胞;图S3)。研究者根据分层聚类(即从GABA1~GABA40,从GLUT1~GLUT48)用数字句柄讨论神经元细胞类型,并在图中细胞类型名称中突出显示1到3个标记基因(如GABA6-PCDH8-ZIC1)。
研究者通过scRNA-seq揭示了金鱼端脑区域内细胞的分子表达谱,以生成细胞图谱。然而,scRNA-seq的组织分离是在失去更精细的空间信息的情况下进行的,空间信息对于定义拓扑同一性和与其他物种的同源性至关重要。因此,研究者沿着端脑的前后轴进行了ST(图1A)。ST(Visium,10x Genomics)是一种基于测序的全转录组原位基因检测方法;在失去细胞分辨率的情况下,该方法保留了mRNA分子的位置信息,每个55μm直径的捕获点排列成一个阵列,中心(X-Y)间隔100μm。研究者从两条金鱼的端脑上采集了8个10μm的冠状切片,每个切片平均间隔100μm(Z)。因此,在像素大小约为100μm时,ST数据包含金鱼端脑中每个基因表达的完整图谱。
为了测试转录组是否能够揭示空间模式并能够描绘金鱼端脑的区域,研究者对采样的6710个Visium捕获点进行了无偏聚类。根据不同的分子图谱获得了17个“空间”簇,并将这些簇映射到了组织切片上(图S4)。尽管聚类过程仅依赖于基因表达,并且基于每个55μm捕获点存在的“大量”细胞,但研究者在每个切片中都发现了明确的空间分割。例如,在前部切片中,浅表斑点与更深的斑点不同。这可能是由于端脑室(室管膜)近端表面存在MFGE8+室管膜胶质细胞(图S3)。更靠后的位置出现了一些更小的、空间上精细的分子区域,类似于解剖学上的细胞核。
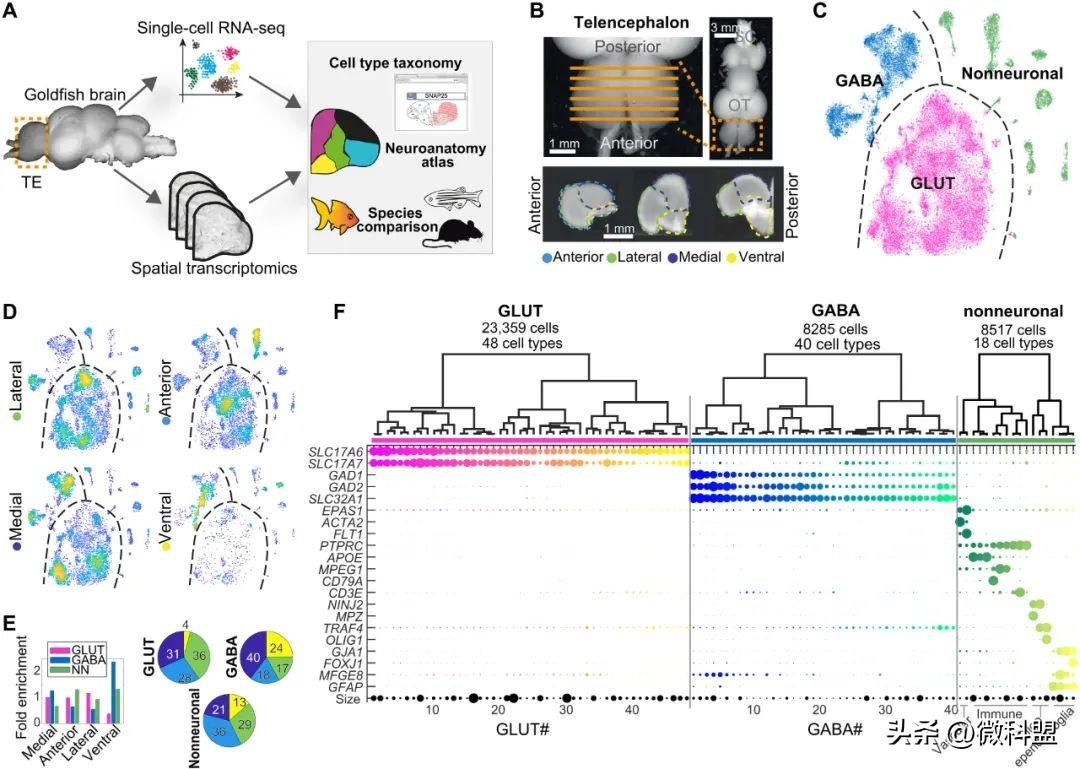
图1.金鱼端脑细胞的组成。(A)本研究的大纲。利用空间转录组学和单细胞转录组学对成年金鱼端脑进行了分析,并将其整合到一个空间定位的细胞类型图谱中,随后进行了物种比较。(B)新鲜解剖的金鱼脑和端脑冠状切片图像,描述了显微解剖的方案。OT:视神经;SC:脊髓。(C)金鱼前脑中所有细胞的t分布随机邻域嵌入(t-SNE)可视化。每个点代表一个细胞,按类别着色。(D)每个显微解剖原点上所有细胞t-SNE的密度散射可视化。黄色,密集;蓝色,稀疏。(E)左图:四个不同解剖面中每个细胞类别占比;可视化为饼状图。右图:不同细胞类型占解剖面总细胞的比值;可视化为条形图。(F)所有细胞类型的聚类树图。底;顶端标记基因表达以点状图可视化。点的大小表示在一个簇中表达每个基因的细胞的百分比;点颜色表示细胞类型。
2 区域分割的神经元类型的空间映射
研究者进一步分析ST如何揭示粗略的解剖分区以证实转录组编码的金鱼端脑中的解剖信息。结合scRNA-seq数据,空间数据集能够定位端脑中分子定义的神经元细胞类型。为了将每种scRNA-seq细胞类型的可能位置映射到解剖位置,研究者应用了一个细胞空间数据集上的类型映射算法。对于每种分子细胞类型,研究者对大脑的前后两个切片中每个点检测到的最高富集基因的表达进行评分和可视化(图S5)。由于ST阵列的每个点可能包含大约10个细胞,因此几种细胞类型可以映射到同一点。接下来,研究者计算每个点的细胞类型得分的加权总和,并使用图1F的细胞类型颜色图可视化88种细胞类型的集成映射(图2A和图S6)。这种图谱可以直观地识别每个点的主要细胞类型,并提供了几个关键信息:(i)分子相关的细胞类型显示出重叠的空间模式,(ii)所有区域都有多种细胞类型,(iii)一些区域与边界清晰的相邻区域明显不同,而其他区域的边界较为分散。
例如,谷氨酸能类型GLUT41~GLUT48在基因表达中构成了一个密切相关的分支,并且都仅局限于前端脑的一个区域(图2A,黄色)。它们与相邻的细胞类型分支(GLUT33~GLUT40)有一些重叠;然而,后者继续延伸到更多的背部和后部。因此,与GABA能簇GABA24~GABA40(绿色)定义的清晰边界相比,前侧区域和更背部区域之间的边界较为分散。
研究者利用视觉颜色边界,在每个端脑视觉部分内手动绘制区域轮廓。描述了基于分子定义的神经元细胞类型的神经解剖学分区。在大多数情况下,研究者按照金鱼神经解剖学常用术语为这些区域定义了名称(图2A,底部)。本研究的解剖分区与传统公认的主要基于细胞结构的分区基本一致。如涉及先前未注释的一个区域[腹侧生长抑素(Vsst),如下所述]以及本研究者发现分子上无法区分的几个注释区域。例如,Dld(背侧区,背侧细分)与更后方的Dp(背侧区)相似,并且与Vd(腹背区)类似,在本研究的注释中没有分为Vs/Vp(腹侧区腹侧上核/腹后核)。虽然这些区别在更高的分辨率或其他上下文中可能仍然相关,但研究者采用了更简化的注释。
3 无偏倚地发现轴向模式基因
接下来,研究者进行了独立细胞类型定位和区域划分,目标是确定在空间分割中最重要的基因。空间限制性基因对物种比较和理解细胞类型的发育起源特别重要。因此,研究者根据限制基因的内外侧和背腹侧轴向范围分析了它们的拓扑分布。
为了获得每个基因的无偏空间评分,研究者计算了沿表达该基因的点的外侧-内侧(X)和背侧-腹侧(Y)坐标的分布。与分散或零星表达的基因相比,编码空间信息的基因定位于特定的区域,没有很分散。此外,就像在X-Y平面上可能存在模式一样,某些基因也可能表现出前后模式。当聚焦于较窄的冠状切片时,可能会忽略这一事实。因此,研究者整合了所有切片和重复的XY空间得分,揭示了沿前后轴的顶级高轴基因的聚合模式。轴向模式不是由单个基因决定的,而是几个基因具有相似的趋势,研究者将其分组为模块。图2B可视化了这个系统的、综合的数据。
研究者发现轴向模式基因表现出同轴相关性;例如,没有基因只有背腹侧,而没有内外侧。相反,模式基因总是在两个或所有(XYZ)轴上表现出轴向富集。此外,研究者观察了所有可能的同轴关系组合。例如,模块3和模块4在前部和外侧以及腹侧(模块3)或背侧(模块4)都富集。神经肽PVALB(模块12)和CARTPT(模块2)分别位于背外侧和腹内侧(图2B和C);神经肽TAC2仅局限于前外侧区域。总体而言,本研究对轴向模式基因的系统评分可以揭示发育起源(形态发生素样),并在设计新遗传体系时为研究人员提供帮助。

图2.金鱼端脑的轴向和区域分割。(A) 基于分子细胞类型的金鱼端脑神经解剖学。顶部:GABA能和谷氨酸能细胞类型的配色方案(树状图)。中部:金鱼的八个冠状切片(金鱼 1),采用Visium ST采样,覆盖整合了所有金鱼端脑细胞类型的加权彩色图。底部:基于上述颜色图差异的区域划分,每种颜色表示不同的区域,以及根据Northcutt建议通过相似区域名称注释的命名法。D,背部区域;V,腹侧区;Dc,Dm的大细胞亚分类;Vsst,腹侧Sst;Ppa,视前核;a,前部;p,后部;d,背部;v,腹侧;m,内侧;l,横向。(B) 金鱼1和2的前几个轴向模式基因沿内外侧(左)和背腹测(右)的SD(每行归一化)热图;点表示根据上面所示的轴向配色方案的空间富集;灰色无点,无富集。右图:每个基因的轴向评分(富集部分的平均值)。(C) 14个轴向模式基因在金鱼端脑中的表达。灰色,低;红色,高。
4 多种GABA能抑制细胞类型
正如最近分析的非哺乳动物,将GABA能细胞鉴定为表达GAD1、GAD2和SLC32A1的细胞,分别编码谷氨酸脱羧酶和GABA囊泡转运蛋白(图1F)。在硬骨鱼中,这些基因被称为皮层下衍生物。与此一致,本研究的ST数据揭示了它们在内侧、腹侧和后部区域富集。在GABA能细胞类型中,观察到了40种细胞类型(图3A),其特征在于特定标记(图3B和C)和组织中的不同位置(图3D)。
首先,GABA能细胞类型在树状图上的层次排列有助于揭示它们的分子相似性,识别联合相关类型的基因,并提供与哺乳动物相似性的线索(图3B和C)。例如,树状图最右侧分支上的细胞类型都表达哺乳动物纹状体标记物MEIS2和PENK。此外,GABA1至GABA6细胞类型共享转录因子(TF)DLX1、NXPH1和ETV1以及prepronociceptin PNO的表达,这些都被称为小鼠皮质中间神经元标记。在小鼠中,中间神经元在新皮质中呈分散分布。将金鱼端脑GABA1至GABA6细胞类型映射到空间数据集,揭示了大细胞背侧分区(Dc)和Dl中类似的分散分布(图3C和D)。
另一个例子是在下一个分支(GABA8~GABA10)中发现的,其与GABA1~GABA6共享CPLX1和ELAVL4的表达。这个相关分支神经肽SST的表达与已知的哺乳动物抑制性中间神经元相似。其中,GABA8和GABA9还表达第二种神经肽NPY(神经肽Y)。然而,与分散的小鼠SST中间神经元不同,GABA8~GABA10主要集中在后端脑的腹中区(图3C和D)。研究者将这个腹侧区域标记为 Vsst,以其在区域模式中最丰富的细胞类型(SST-NPY)命名(图2A)。然而,其他标记特定小鼠间神经元类型(Pvalb和Vip)的显著标记在金鱼端脑中表现完全不同:它们不是特定标记中间神经元(GABA1~GABA10),甚至不是GABA能类神经元的。例如,唯一的GABA能VIP(GABA30)不表达其他局部中间神经元。这种细胞类型位于Vd区(图3B以及图S5和S9),研究者发现该区域富集类似于抑制性投射类的神经元[中等多棘神经元(MSN)]。此外,在成年金鱼中,神经节隆起衍生的中间神经元的发育性大脑皮层下分裂是矛盾的。在哺乳动物中描述的许多区别标记物[例如内侧神经节隆起(MGE):Sst和Pvalb;尾神经节隆起(CGE):Vip、Lamp5和Sncg]显示重叠表达(图S8)。
GABA能类神经元GABA11~GABA40沿中线分布,其中GABA11~GABA22在腹侧区(Vv)的腹核中富集,GABA24~GABA40在背侧(Vd)富集。其中,一小群(GABA15)类似于哺乳动物多巴胺能神经元:它们在小鼠中特异性表达酪氨酸羟化酶(TH)、多巴胺囊泡转运蛋白(SLC6A3)和泛中间神经元TF SP8标记基因。在空间上,这种细胞类型出现在一个微小的腹侧内侧核中(图3B和图S5),与斑马鱼端脑中描述的th1表达非常相似。
最后的Vd分支(GABA24~GABA40)由几个重要的标记物标记,包括TF SIX3和MEIS2、神经肽PENK和多巴胺受体DRD2(但不包括DRD1),这些标记在小鼠纹状体中MSN的GABA能类中富集。这就提出了哺乳动物纹状体和这些特别多样化和离散的金鱼类型之间可能的类比。最后,研究者鉴定了沿着中线的脑室周围区推定的GABA23 GABA能神经母细胞。正如对鱼类和蝾螈中成体神经母细胞的描述,该群体特异性表达MEX3A和TUBB5。
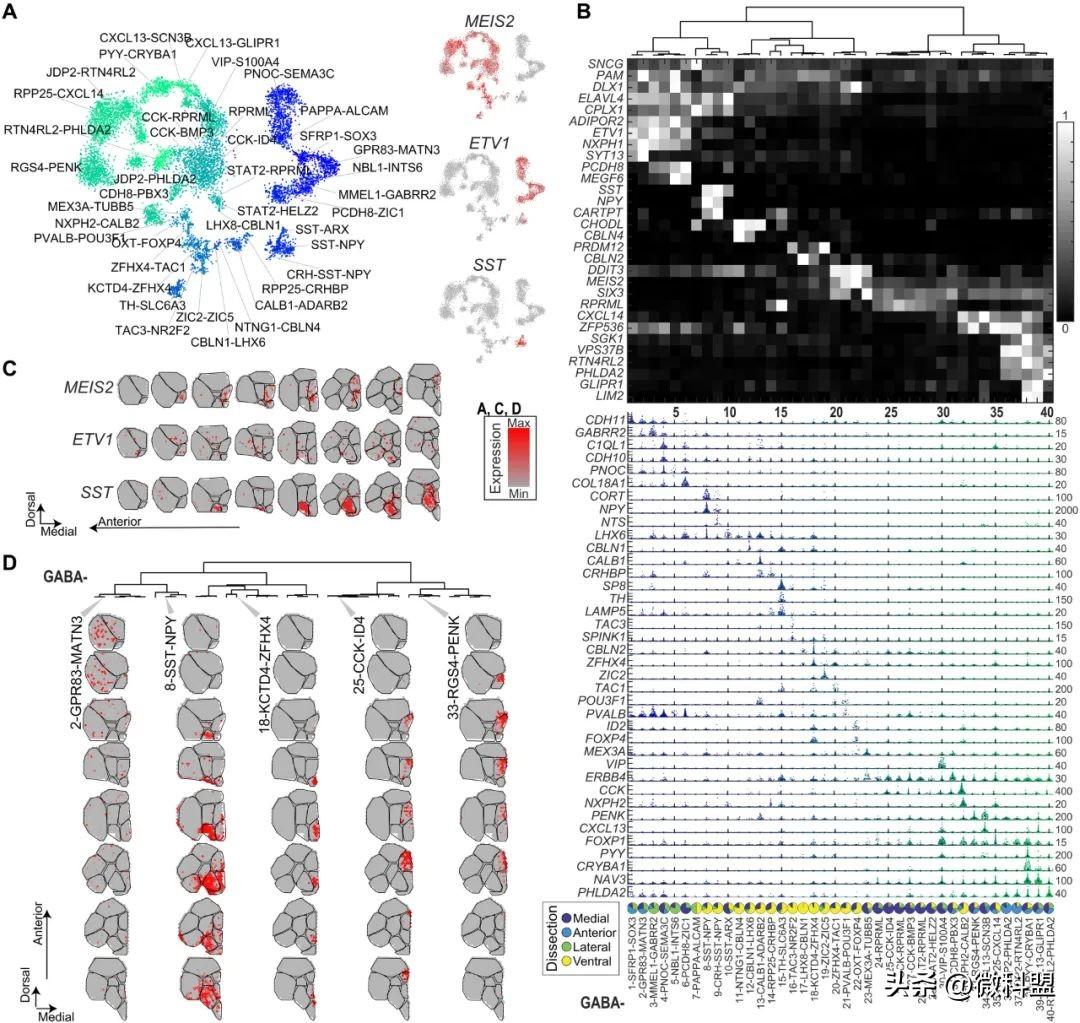
图3.金鱼端脑中的GABA能神经元。(A) 金鱼前脑中GABA能神经元的t-SNE可视化。每个点代表一个细胞,根据细胞类型配色。右图:三种神经元标记基因的表达。(B) 所有GABA能类神经元均按树状图顺序排列(GABA1至GABA40),最重要标记基因表达可视化为热图(白色,高;黑色,低)。中部:小提琴图,其中每个点代表一个细胞;最大表达量(UMI)如右侧所示。底部:四种显微解剖中每种细胞类型的占比,以饼图形式可视化。(C) 三种神经元标记基因的表达[如(A)]以及ST中八个前后端脑冠状半球切片的表达。(D) 整个GABA能树枝状图中Visium点的空间相关性示例:八个a-p冠状切片(行)中的五种scRNA-seq细胞类型(列)。
5 谷氨酸能细胞类型及其空间分布
研究者发现了48种分子定义的谷氨酸兴奋性细胞类型(图4A和B),具有独特、明显的空间分布(图4C和图S5)。它们是通过谷氨酸囊泡转运蛋白SLC17A6(VGLUT2)的表达来定义的。这些类型中超过一半还表达SLC17A7(VGLUT1)(图1E)。这与小鼠形成鲜明对比,在小鼠中,Slc17a6几乎完全保留在间脑,Slc17a7在前脑中占主导地位,并且只有少数细胞类型共表达两种转运蛋白(例如,皮质L5、脾后皮质和一些杏仁核)。在金鱼中,SLC17A6+ SLC17A7+簇存在于谷氨酸能表达的所有区域,但它们在Dl(aD1)和Dld前部表达较少。然而,金鱼端脑的谷氨酸能神经元也被TBR1标记,TBR1是一种TF,在成年小鼠中标记皮质锥体神经元,并在较小程度上标记皮质锥体下神经。谷氨酸能细胞类型在端脑背侧和外侧的大脑皮层区域丰富(例如,Dm、Dc和Dl),与在大脑皮层下区域(例如,Vv和Vd)高度丰富的GABA能细胞几乎具有互补的空间模式(图S5和S6)。
谷氨酸能神经元的第一次层次划分反映在基因表达和空间分布上:reelin(RELN)、大麻素受体1(CNR1)和甲状腺激素核受体(NR2F2)定义了相关细胞类型的分支,分别是树状图的右侧(GLUT20~GLUT48:RELN和CNR1)和左侧(GLUT1~GLUT19:NR2F2)(图4B和C)具有高度特异性。这些基因还表明不同的空间模式,几乎没有相互重叠:CNR1位于前面且位于Dm区域,而NR2F2位于Dl和Dm区域的后面(图4C)。NR2F2型GLUT1~GLUT19被细分为五个不同的组(图4B)。例如,GLUT1~GLUT6表达BCL11B并占据腹侧D1(图4B至D,以及图S5)。GLUT7~GLUT9的空间模式位于金鱼端脑Dm和Dc的背部(图S5)。树状图的第二个分支(CNR1 GLUT20~GLUT48)被分为六个不同的组(图4B)。例如,GLUT29~GLUT32表达NEUROD1和NOCT并在前Dm中发现;GLUT41~GLUT48表达EOMES并位于aD1中(图4B至D,以及图S5)。
GLUT14和GLUT15位于后端脑的最腹侧部分(图S5)并表达TF OTP和三肽激素TRH。在小鼠中,这些基因准确地标记了小鼠下丘脑和纹状体杏仁核中的几个兴奋性肽能神经元。最后,研究者观察了谷氨酸能神经母细胞群:GLUT26表达MEX3A和TUBB5,与GABA能神经母细胞(GABA23)相似。与仅限于中线的GABA23不同,谷氨酸能神经母细胞排列在端脑室管膜的整个范围内(图S5)。这种模式特别类似于金鱼室管膜胶质细胞的分布(例如,HES5+和MFGE8+;见图S3),它们最接近端脑室。星形胶质细胞相关的HES5+放射状胶质细胞在小鼠中充当成体神经源性干细胞。因此,这些细胞类型可能共同构成了成年金鱼端脑的脑室周围神经源性生态位。

图4.金鱼端脑中的谷氨酸能神经元。(A) 金鱼前脑谷氨酸能神经元的t-SNE可视化。每个点代表一个细胞,根据细胞类型配色。(B) 所有谷氨酸能类型,按树状图顺序(GLUT1~GLUT48),最重要标记基因表达可视化为热图(白色,高;黑色,低)。中:小提琴图,其中每个点代表一个细胞;最大表达量(UMI)如右侧所示。底部:四种显微解剖中每种细胞类型的占比,以饼图形式可视化。(C) 两个标记基因NR2F2和CNR1的表达,在t-SNE[如(A)]和ST中的八个前后端脑冠状半球切片上可视化。(D) Visium空间相关性的谷氨酸能树状图示例:八个前后冠状切片(行)的四种scRNA-seq细胞类型(列)。
6 硬骨鱼端脑分区的分子特征
研究者对轴向模式基因(图2)和分子细胞类型图谱(图3和图4)进行了分析,从而在分子上确定了区域边界。为了便于未来的功能探索,研究者提出了相反的问题:哪些基因(图5A和B)和细胞类型(图5C)能最好的定义每个区室?例如,上述SST主导的Vsst也由SST受体结合皮质抑素(CORT)定义。TF eomesodermin EOMES(也称为TBR2)巧妙地标记了aD1。钙结合NECAB2是腺苷2A和mGluR5受体的调节剂,不仅集中在背侧区(Dlv)的腹侧亚区中,而且还分散在整个pDm等区域中。相比之下,白质为主的DC确实可以用髓磷脂结合蛋白MBP来描述,并且在少突细胞群中富集。总之,这种分析精确地描述了与已知解剖学描述一致的基因的分子基础。

图5.区域特异性标记基因和细胞类型。(A) 通过ST鉴定的最富集的区域特异性标记基因,以点图表示:圆圈大小代表每个区域中每个基因的标准化平均表达。(B) 来自(A)的基因的代表性实例,具有跨八个前后端脑部分的表达空间分布。灰色,低;红色,高。(C) 细胞类型到区域的富集显示为点图,其中点大小表示细胞类型表达谱和区域表达谱之间的相关性。(D) 在ST和HCR-FISH的匹配切片中,验证沿端脑前后轴的NEUROD6表达区域的转换。HCR的切片概述分段显示,下面是标准化的表达量表。比例尺,0.5 mm(ST和HCR概述)。(E) 仅两组谷氨酸簇的加权空间细胞类型富集(绿色,GLUT20,GLUT21,GLUT30和GLUT31;红色,GLUT5,GLUT12,GLUT16和GLUT18),表明两组的Dm-Dl区域转换会导致3D弯曲/角。
7 “扭曲”轴向分区的兴奋性细胞类型
本研究分析中的多个数据点强调了沿完整前后轴的分子表征的重要性,以便更好地概括三维(3D)的基础结构。例如,研究者发现Dm和D1区域前部和后部大脑皮层之间基因表达和细胞类型组成发生了有趣的变化。首先,研究者注意到NEUROD6,一个已知标记哺乳动物海马锥体神经元的基因,在前部(aDm和Dm)位于背内侧,但在后部,似乎将区域“切换”到Dld [对应于Northcutt Dp](图2C和5D)。值得注意的是,神经肽PVALB似乎做出了相反的转换,在更前面的地方,它位于背外侧(Dl/pDl),但位于背内侧区域(pDm)的后面(图2C)。这种趋势反映在与PVALB和NEUROD6共享轴向模式表达的其他基因(例如 PVALB、CAMK2A、EFNA1、SERPINE1和EMID1)(图2B和5B)。几种谷氨酸能细胞类型的空间分布也支持了这一观察结果:沿着前后轴,GLUT20,GLUT21,GLUT30和 GLUT31从Dm切换到Dld,而GLUT5、GLUT12、GLUT16和GLUT18从D1切换到pDm(图5E 和图S5)。
因此,单纯基于轴向分裂的分区并不总是与基于细胞类型的模式一致。如果基因表达和细胞类型组成代表功能,那么这里描述的内侧-外侧Dm-Dl转换值值得关注:研究者认为,两组神经元群可能代表了不同的功能单元,分别由NEUROD6(Dm-Dld)和PVALB(Dl-pDm)描述。。在三维空间中,每组都会形成一个弯曲或角,类似于啮齿动物海马体的3D形状。这些弯曲在一起看起来会相互缠绕,就像两个分子和功能不同的区域的前后扭曲。
8 金鱼和小鼠细胞类型的分子相似性
不同脊椎动物的前脑结构各不相同。如今,小鼠大脑对于我们了解脊椎动物神经系统至关重要,但它在大约4.5亿年前就与硬骨鱼产生了分歧。对两栖动物、胡须龙和海龟的端脑的比较研究揭示了高度保守的细胞类型,即使与小鼠大脑相比也是如此。鉴于金鱼和小鼠之间巨大的进化差距,识别分子相似性和同源大脑区域可能具有挑战性,但可能有助于更好地了解脊椎动物大脑结构的进化。因此,研究者继续对金鱼端脑进行研究,并与小鼠前脑进行系统比较。简而言之,研究者将金鱼分类法与已发表的小鼠端脑细胞类型分类法相结合。为此,将物种的数据集合并到非神经元细胞、GABA能神经元和GLUT能神经元的混合物种数据集。对于每个细胞类别分别进行特征选择和降维,并迭代数据集成工具Harmony,直到两个物种的细胞图谱的嵌入收敛(S7)。然后,应用了两种并行方法来帮助比较分析:(i)对混合物种伪细胞类型进行聚类,以及(ii)金鱼细胞与小鼠细胞类型进行KNN(k-近邻)分类器(图6A)。
为了避免每种方法固有的偏差(例如簇大小依赖性),研究者提供了两种方法之间一致的细胞类型示例,即两种方法都确定了小鼠与金鱼的相似性(图6和图S7)。然后,检查了这些保守神经元细胞类型的空间图谱,以评估空间范围或大脑区域可能在多大程度上被保守。
9 神经胶质细胞的保护和广泛的金鱼免疫生态位
非神经元细胞在两个物种之间表现出总体上很大程度的保守性,但也有一些有趣的例外。在神经胶质细胞群中,本研究检测到三种类型的室管膜神经胶质细胞,它们都与不同的小鼠星形胶质细胞群密切相关(图6B和C,以及图S7)。 他们表达了一系列已知的哺乳动物星形胶质细胞标记,包括MFGE8、GFAP和GJA1(图S3)。与硬骨鱼和哺乳动物对应物之间的功能差异一致,金鱼室管膜神经胶质细胞表达的基因通常在小鼠星形胶质细胞中检测不到,例如金属硫蛋白MT4、δ样缺口配体DLL1、单羧酸转运蛋白SLC16A9和分泌糖蛋白SLIT2(在发育中具有轴突引导功能)(图S3)。此外,与哺乳动物不同,所有种群几乎完全局限于软脑膜和脑室表面(图S3)。表达小鼠放射状胶质细胞标记物HES5(室管膜胶质细胞-HES5-DLL1)的群体确实与成年小鼠的两个神经源性生态位、齿状回和室下区的放射状胶质细胞群体密切相关(图6B)。不同物种之间的室管膜细胞的基因表达和沿脑室的分布都非常密切(图6B以及图S3和S7)。鉴于它们在脑脊液循环中必要的保守功能以及这种专门化所需的基因产物,这是预料之中的。
金鱼少突胶质细胞生态位重建了成年小鼠大脑中的整个谱系,从少突胶质细胞祖细胞(OPC)到定向少突胶质细胞(COP),最后是成熟的髓鞘少突胶质细胞(图6B和图S3和S7)。这一观察结果既证实了其保守的发育起源,也证实了与室管膜细胞类似的髓鞘形成中保守的特殊功能。然而,金鱼少突胶质细胞也表达CLDN19、MPZ和SPARC等基因(图S3),这些基因在小鼠中完全保留给外周神经系统的髓鞘细胞即雪旺细胞。鉴于哺乳动物雪旺细胞的发育起源于神经嵴,而不是神经管,这一点尤其值得注意。然而,由于小鼠数据集中包含少量雪旺细胞,本研究的分析中的两种方法都忽略了小鼠雪旺细胞和金鱼少突胶质细胞之间的这种意想不到的关系(图6B和C,以及图S7)。
与小鼠相比,金鱼端脑免疫生态位(以PTPRC(CD45)为标志)非常多样化(图S3)。与小鼠大脑类似,本研究检测到了几种类型的巨噬细胞(APOE)和小胶质细胞(MPEG1)(图6B和C,以及图S3和S7)。一些免疫细胞与小鼠大脑中通常未检测到的外周免疫系统的骨髓和淋巴成分相似,例如粒细胞或单核细胞(免疫-CCL2-CCL24和免疫-AQP1-AQP9)、B细胞(免疫-CD79B)、和T细胞(免疫-CD3E-SLA2)。因此,与带注释的小鼠前脑数据集的相似性可能会产生误导(图6B和C以及图S7)。例如,粒细胞样免疫AQP1-AQP9在KNN分析中类似于室管膜(EPMB)和下连膜(HYPEND)细胞,在聚类方法中类似于星形胶质细胞(Bergmann胶质细胞)。假定的T细胞免疫-CD3E-SLA2类似于脉管系统、(VLMC/ABC)和上皮细胞(脉络丛)。然而,人们很容易推测,这一观察结果是否表明小鼠和金鱼细胞类型进化的功能相似。
10 SST中间神经元的分子相似性和空间相异性
在GABA能类中,物种比较检测到潜在的保守细胞类型,两个物种中都有分子相似的对应物(图6D到H)。首先,KNN和集成聚类分析都证实了中间神经元(GABA1~GABA10)和假定的投射(类似MSN)抑制类型(GABA16~GABA40)之间的总体划分。
GABA1~GABA7类似于一组皮质和海马中间神经元(TEINH14~TEINH18),是CGE-和MGE-衍生的表型(图6D)。金鱼SST簇GABA8~GABA10是高度保守神经元类型的一个特别值得注意的例子。在整合聚类KNN分析中,GABA8~GABA10的评分与小鼠皮层SST中间神经元TEINH19和TEINH21的评分非常接近(图6D和图S7)。然而,如上所述,这种神经元类型的空间分布在两个物种中有所不同(图6E):在小鼠中,SST中间神经元以分散的模式分布在整个端脑。在金鱼中,SST类型GABA8~GABA10的分布集中在大脑皮层下的腹侧部分,这个区域称为Vsst(图2A和图S6)。ST和荧光原位杂交(FISH)均显示大脑皮层中稀疏的SST表达,并证实了研究者对大脑皮层下观察到致密的SST+核(图6E和7以及图S9)。由于它们的聚集空间模式,研究者推测SST类型可能更接近于不同的皮层下类型,例如哺乳动物纹状体杏仁核的Sst+种群。研究者将所有GABA能群体与另一个小鼠中央杏仁核数据集整合(图S8)。然而,金鱼-SST群体GABA8~GABA10与小鼠中央杏仁核表达Sst的投射类型无关(图S8)。相反,它们与杏仁核Sst+ MGE-中间神经元群体紧密聚集在一起,并且在GABA10的情况下,与内侧杏仁核的一个Sst+群体紧密聚集,所有这些都共享调节迁移的 TF Nkx2-1的表达(图S8)。总之,SST中间神经元是小鼠皮质和皮质下SST中间神经元高度分子保守细胞类型的有力例子。然而,由于它们不同的定位,需要进一步的工作来阐明这些条件是否意味着不同的功能。
同样的分析还揭示了金鱼中间神经元群体对于神经节隆起的发育起源更矛盾的态度(图S8)。总体而言,两个神经元间分支GABA1~GABA7和GABA8~GABA10分别与CGE和MGE紧密映射。此外,GABA8~GABA10 SST中间神经元不仅表达MGE相关神经肽Y(NPY)和TF LHX6,还表达CGE相关基因SNCG、LAMP5和ADAR2B以及TF SP8和NR2F2。类似地,GABA1~GABA7不仅表达CGE相关的TF PROX1、NFIB和NFIX,而且还表达MGE相关的NXPH1、NOS1和PVALB。此外,虽然保守转录因子的整合嵌入和表达所揭示的整体相似性可能更好地反映了共同的进化,但四足动物[例如蝾螈和蜥蜴]中的几个图谱揭示了与哺乳动物中间神经元类明显更高的分子相似性。

图6.金鱼和小鼠端脑细胞类型的跨物种比较。(A) 金鱼和小鼠端脑之间的比较物种分析方案。 每个细胞类别都会集成两个物种的数据集(Harmony),然后使用噪声(DBSCAN)聚类或KNN分类器对应用程序进行基于密度的空间聚类。(B) 以热图形式显示的非神经元细胞种类比较与概述的保守细胞类型(KNN 分类器)。(C) 非神经元细胞种类比较可视化为t-SNE。点代表细胞,根据金鱼细胞类型着色。灰色,小鼠细胞;灰色轮廓,整合集群(见图S7);粉色轮廓和标签,小鼠细胞类;突出显示小鼠雪旺细胞。(D) GABA类物种比较(KNN分类器),其中突出显示了与整合簇的一致性的示例。(E) SST中间神经元类型的分布,与金鱼ST和小鼠ISH的空间相关性,放大到同皮质中分散的SST表达(Allen小鼠大脑ISH 图谱)。(F) 通过与金鱼ST(左)和小鼠ISH(右)的空间相关性来分布(假定的)纹状体神经元类型。(G) 两种小鼠纹状体标记物在金鱼ST(左)和小鼠ISH(右)中的表达。(H) 通过金鱼ST(左)和小鼠ISH(右)的空间相关性来分布(假定的)苍白球神经元类型。(I) GLUT能类物种比较(KNN分类器),突出显示了与集成簇一致性的示例。(J) 通过与金鱼ST(左)和小鼠ISH(右)的空间相关性来(假定的)海马后神经元类型。(K) 金鱼ST和小鼠ISH中五个基因在(假定的)的海马下/海马后形成中的表达。
11 金鱼Vd中的纹状体和苍白球衍生物
金鱼GABA能细胞类型的另一个分支(GABA16~GABA40)总体上与局部抑制性皮质中间神经元类型的相似性较低(图6D)。相反,小鼠中一些最相关的细胞类型包括纹状体中型基棘细胞以及小鼠隔膜和苍白球的抑制性神经元。例如,金鱼中的GABA25~GABA28与小鼠MSN分组一致(图6F)。ST将这些细胞映射到Vd内侧(图2A,紫色,图S5)。此外,小鼠纹状体标记物PENK和MEIS2仅在金鱼Vd中聚集,形成一个连续区域实体,类似于小鼠纹状体的情况(图6G)。最后,物种比较一致地将GABA33~GABA39与小鼠隔膜和苍白球抑制细胞类型归为同一类,最显著的是与终纹床核(BST)群体DEINH5(图6D和图S7)一致。这些细胞类型位于Vd的更前面(图6H)。总之,这些分子组成和空间分布的相似性表明金鱼与小鼠脑核的Vd具有同源性。本研究的分析进一步表明它们分别解析为Vd最前面和后面的苍白球和纹状体样区域(包括传统上称为Vs和Vp的区域)。
12 金鱼Dlv中海马后形成的同源物
与GABA能细胞类型(当然还有非神经元类型)相反,谷氨酸能细胞类型在本研究中的两种物种比较方法之间表现出较低的一致性。这意味着兴奋性细胞类别的保守性较低(图6I至K),正如爬行动物大脑皮层的描述一样。
尽管如此,研究者发现GLUT20~GLUT22和GLUT37~GLUT40可能与海马神经元相关:无论采用何种分析方法,这两组都与小鼠海马相似(图6I)。然而,并不能精确定位到单个小鼠细胞类型。相反,根据方法的不同,它们的关系在小鼠海马分区之间交替变换[例如,TEGLU24(CA1)、TEGLU23(CA3)和DGGRC1-2(DG)](图6I、图S7和表S1)。金鱼和小鼠中这些细胞类型之间的相似性可能部分是由小鼠海马富含的基因C1QL1和NEUROD6来解释的,这些基因在GLUT21和GLUT22中高表达。两组(GLUT20~GLUT22和GLUT37~GLUT40)分散分布在Dm和D1区域(图S5)。总之,这些发现表明,与小鼠相比,金鱼的分子保守性有限,并且空间上定义的海马结构较少。
在谷氨酸能神经元中,最为一致的是一组金鱼神经元GLUT1和GLUT2,其得分与海马下托谷氨酸能细胞类型(TEGLU14)的得分非常接近(图6I和J,以及图S7)。金鱼细胞类型对应映射到分子上不同的侧腹区域点Dlv(图6J和图S5)。CBLN1、CBLN2、NWD2、ETV1和RSPO2的共同表达体现了物种间的相似性(图6K)。此外,KNN分析仅表明小鼠海马内嗅和后海马神经元类型(TEGLU5和TEGLU21)密切相关(图6I)。在小鼠中,内嗅区和下托属于海马后结构。它们在海马-皮质相互作用和空间导航(例如头部方向)中发挥主要作用。总之,金鱼中这些假定的下托或海马后形成样神经元表现出保守的分子特征,同时集中在一个区域(Dlv),根据硬骨动物外翻端脑的发育,该区域在拓扑上也与海马后结构相对应。
13 金鱼和斑马鱼的硬骨端脑同源性
物种之间同源物的识别依赖于进化拓扑图的识别,因此对物种发展的详细了解极大地帮助了物种同源物的识别。金鱼和斑马鱼等硬骨鱼具有外翻的端脑结构。然而,斑马鱼(Danio rerio)是研究最深入的硬骨动物的代表,并以拥有大量研究工具而闻名。斑马鱼的分子、发育和连接性数据表明,斑马鱼端脑的拓扑结构可以与哺乳动物的情况进行比较。
因此,接下来研究者将成年斑马鱼端脑细胞类型调查的数据与金鱼端脑进行整合。在这个分析中,研究者利用了相对更好的斑马鱼转录组注释,并用它来注释金鱼基因,尽管全基因组复制产生的旁系同源物再次被折叠。将斑马鱼端脑的21种神经元类型与金鱼端脑的88种神经元类型进行比较后,与小鼠(图6和图S7)相比,这些端脑物种之间的细胞分类高度保守(图7A和C)。正如预期的那样,Pandey 等人注释的大脑皮质或皮质下细胞类型在很大程度上是根据神经递质类别划分的,它们分别主要是谷氨酸能或GABA能。
在细胞类型水平上,Subpallial_01对应于几个金鱼中间神经元群,包括金鱼Vsst中聚集的Sst表达类型(图7D和E,SST)。作者描述了斑马鱼中间神经元类中类似的VIP缺失,并得出结论,在斑马鱼中没有与CGE衍生的哺乳动物中间神经元相对应的群体。中等棘状金鱼细胞GABA23~GABA40类似于斑马鱼细胞群Subpallium_02和Subpallium_05(图7D和E,PENK)。最后,斑马鱼群(Pallium_02)与假定的金鱼海马后细胞类型GLUT1~GLUT3的同源物非常相似(图7D和E,CBLN1)。正如本研究的数据所示,作者将这个群体定位在Dlv中,并独立报告了与小鼠海马下托相同的分子相似性。尽管采样深度、注释和空间映射阻碍了更详细的分析,但这一比较证实了金鱼端脑的分子细胞类型分类可以为其他端骨动物的研究提供便利。

图7.硬骨端脑细胞类型的跨物种比较。(A和B)金鱼和斑马端脑的t-SNE可视化比较物种分析,并突出显示每个物种的细胞类型;(A)金鱼和(B)斑马鱼。每个细胞类别,两个物种的数据集被整合(Harmony),然后进行DBSCAN聚类。(C)每个细胞类型比较,使用KNN分类器评分。(D) CBLN1、PENK和SST在整合的硬骨数据集中的表达; 按物种来源着色的点(细胞)。(E)利用HCR-FISH验证ST中检测到的基因表达。上行:Visium(左)和HCR(右)中检测到的基因的相应部分概述,其中每个点代表一个分段的荧光细胞,通过标准化表达着色。下行:放大的原始荧光信号,如概述部分所示。
讨论
本研究将ST与scRNA-seq相结合,生成了金鱼端脑的神经元细胞类型图谱。金鱼是一种类似鲤鱼的硬骨鱼,具有向外生长(外翻)的端脑特征,与射线鳍鱼相似。由此产生的数据集将为分析脊椎动物的神经元类型进化提供关键资源,并提供了比较哺乳动物、爬行动物和两栖动物的各种新兴前脑细胞类型图谱的庞大数据集。
本研究发现了88种GABA能和谷氨酸能神经元类型以及精确的空间映射。研究者发现,在其他物种中存在的所有主要种类的前脑细胞类型也存在于金鱼中。此外,在与小鼠端脑的系统比较中,许多神经元群在小鼠中有直接对应物,其中一些指向保守的大脑区域。
ST揭示了数千个基因的表达模式。本研究介绍了一种对端脑轴向模式基因表达进行空间评分的有效方法。该方法有助于系统地识别具有空间限制模式的基因,从而了解潜在的区域特异性范围和解剖学轮廓。在本研究的例子中,它确定了中外侧轴和背腹轴中最有潜力的标记。许多这些模式形成基因在所有三个轴上都富集,因此符合当前分子不同区域或细胞核的解剖学轮廓。这些空间信息基因也可能为脊椎动物大脑结构的潜在发育过程和进化起源提供线索。例如,后续研究可以使用追踪方法来追踪在成人端脑中保留空间信息的TF。
除了识别单个基因的空间限制表达之外,本研究还使用ST将所有细胞类型映射到它们的位置。假设分子定义的神经元类型代表一个特定的功能单元,然后对所有细胞类型的组合图进行分割,以提出金鱼端脑的区域神经解剖学。此外,虽然这种方法通常与传统的基于细胞结构的端脑图谱一致,但本研究的分区揭示了分子-神经元,因此可能是区域划分的功能基础。这一推测得到了几种细胞类型的支持,这些细胞类型在研究者对金鱼细胞类型与小鼠端脑分类的系统比较中具有分子相似性。在这里,金鱼端脑内的一些分子相似的神经元类型暗示了与小鼠端脑的同源性:它们在拓扑上与小鼠前脑中的对应位置相似的位置上显示出离散的空间分布。例如,本研究的分析表明,金鱼Vd中的细胞类型与小鼠纹状体结构域具有假定的同源性。本研究鉴定了小鼠MSN和金鱼的GABA26-GABA28型GABA能细胞的分子相似性,并鉴定了斑马鱼的白质下对应的GABA26-GABA28型。MSN样金鱼种群和纹状体标记物PENK沿着Vd的前后范围特异性地定位到金鱼的皮层下的限制区域。这进一步符合Vd与纹状体本身的拓扑对应关系。
此外,金鱼的背外侧区(Dlv)的腹侧部分有一个假定的与小鼠海马后区形成的同源物,特别是下托。这是基于小鼠海马后细胞类型与金鱼细胞类型GLUT1至GLUT3之间的分子关系。先前的病变研究表明Dlv与空间认知有关,并且在啮齿动物的下托和金鱼的Dlv中都发现了头方向选择细胞。这反映了这些区域的分子和功能水平之间有趣的对应关系,并且根据外翻-外翻的发育模式在拓扑结构上是一致的。最近在蝾螈的背内侧大脑皮层中发现了与哺乳动物海马后结构类似的结构,除了共有的分子标志外,中尺度连接也类似于爬行动物和哺乳动物。
另一方面,与海马结构的其他部分的相似性差异较小。NEUROD6在小鼠海马CA1-2-3中表达最强,映射到假定的角状Dm-Dld区域。此外,在物种比较中,研究者发现了小鼠海马固有神经元与几种金鱼细胞类型的相似性,这些细胞类型分散在Dl和Dm区域。它们都被认为是硬骨鱼海马的同源物。研究者推测,结合这些功能上的争议,本研究的分子研究结果表明,与哺乳动物相比,空间行为的区域界限更少。本研究结果为这一知识体系提供了一个分子轴,这可能有助于未来的功能表征。研究者预计,多模块方法,如遗传策略与功能记录和连接图谱的结合使用将带来决定性的见解。
在空间背景下对细胞类型映射的综合分析也有助于研究者发现假定的前后神经解剖学:从前向后,两组分子相关的细胞类型从Dm-Dld和Dl-pDm过渡。在纯细胞结构方法的基础上,这可能会被忽略,并需要更精细的空间映射。对于这种结构观察的进一步验证,理想情况下需要一种更无缝的、更完全的方法,能够使足够多的基因来检测多种相关的细胞类型。由于每个细胞类群也明显地以NEUROD6(Dm-Dld)和PVALB (Dl-pDm)的表达为标志,这有助于它们的初步功能探索。例如,人们很容易推测,这些区室的扭曲结构是否导致了上文讨论的关于海马结构功能上的金鱼对应位置的争议。研究者建议新的证据需要考虑到精确的前后对齐,理想情况下,将功能测定与遗传标记结合起来,如上述例子中的NEUROD6(小鼠海马标记)和PVALB。
然而,并不是所有的分子相似性都是同源性的明确例子。例如,多巴胺能细胞类型GABA15(TH+ SLC6A3+)与小鼠嗅球的多巴胺能群相关(图6D和图S7),但其他多巴胺能群(例如小鼠中脑)没有进行比较。Th1在斑马鱼中也有类似的位置,但与斑马鱼不同的是,金鱼的端脑与嗅球是分开的。因此,这些多巴胺能类型的相似性是没有共有的功能或共有的拓扑支持的分子趋同的潜在情况。与斑马鱼一样,本研究还描述了许多基因表达的不一致,这些基因标志着小鼠和其他四足动物MGE和CGE中中间神经元的发育生态位。
在对比小鼠的一个特别值得注意的例子中,表达神经肽SST的中间神经元群形成了一个完全同质的腹侧核,将其标记为Vsst。小鼠前脑的SST神经元作为皮层和海马锥体细胞的抑制性中间神经元。然而,金鱼的核样聚集进一步挑战了它们作为局部抑制细胞的小鼠功能。因此,Vsst和其他SST细胞可能对进一步探索具有重要意义,以揭示这种独特的细胞类型在功能和行为上的进化保守程度。
通过绘制金鱼和小鼠之间的细胞类型图谱,发现了这两个物种大脑区域可能存在的分子同源性。尽管它们的进化距离遥远,但关于小鼠神经解剖学和细胞类型的大量可用信息使本研究能够将金鱼的发现置于更广泛的神经行为学背景下。这使本研究能够明确在大脑进化过程中保留下来的关键成分。请注意,这些信息当然不能完全描述同源性:本研究没有遵循每个区域的发育和进化路径,并且将细胞类型与区域连通性联系起来的关键内容需要共同的努力。然而,本研究的方法不仅测试了单独的、选择的标记基因的表达,还测试了代表金鱼端脑的功能成分的分子定义的细胞类型。
总的来说,本研究提供了对硬骨动物端脑的细胞特征和转录结构的深入分析,以揭示与哺乳脊椎动物前脑的神经进化相似性。研究者相信,结合神经解剖图谱和分子细胞类型信息,将有助于更深入地表征这些区域的功能。然而,本研究的数据集并不是详尽的,技术的持续发展将在未来产生更深入、更好的空间分辨率的硬骨鱼图谱。此外,由于额外的基因组复制而产生的金鱼旁系同源物的作用可能在未来得到更彻底的探究,并为新功能化或亚功能化的进化提供重要的视角。
为了促进未来对金鱼的研究,本研究建立了一个可访问的网络资源,用于自由探索不同细胞类型和空间背景下的基因表达。这些数据和资源可能对金鱼、斑马鱼和硬骨鱼研究具有价值,并能够在脊椎动物比较神经科学中开辟其他领域。例如,在行为任务期间,特定种群可能会在基因和/或空间上进行记录。在这种情况下,金鱼是特别令人感兴趣的,不仅仅是因为它的体型很大,而且它易于实验操作。金鱼现在分布在世界各地的自然水域,这可以进一步促进将实验获得的数据与自然水体中的真实行为进行比较。本研究金鱼的神经生物学不仅揭示了它们非凡的能力,还可以增强了对超越物种界限的基本大脑功能的理解。



















 鲁公网安备37020202370234号
鲁公网安备37020202370234号